
وکتورهای کلونینگ برای یوکاریوت ها – بخش سوم (وکتورهای کلونینگ برای گیاهان عالی)
✍ وکتورهای کلونینگ برای گیاهان عالی
توسعه وکتورهای کلونینگ برای گیاهان عالی در دهه ۱۹۸۰ صورت گرفت به طوری که استفاده از آن ها منجر به تولید محصولات زراعی تغییر یافته ژنتیکی (GM) شد .
آگروباکتریوم تومه فاشینس – به عنوان کوچکترین مهندس ژنتیک طبیعت
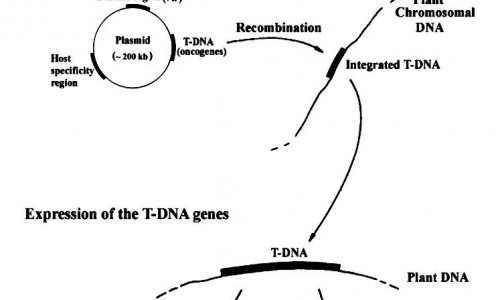
اگرچه پلاسمیدهای طبیعی در گیاهان عالی شناسایی نشده اند، یک پلاسمید باکتریایی به نام Ti ، حاصل از آگروباکتریوم تومه فاشینس، دارای اهمیت زیادی است.
آگروباکتریوم تومه فاشینس یک میکروارگانیسم ساکن خاک است که در بسیاری از گیاهان دولپه ای ایجاد بیماری گال تاجی می کند. بیماری گال تاجی زمانی ایجاد می شود که تهاجم باکتری آگروباکتریوم تومه فاشینس از طریق زخم روی ساقه گیاه صورت بگیرد، که پس از آن باکتری باعث تکثیر سرطانی ساقه در محل تاج می شود.
توانایی باکتری آگروباکتریوم تومه فاشینس مربوط به حضور پلاسمید Ti ( القا کننده تومور) در سلول هایش می باشد. این پلاسمید اندازه ای بزرگ تر از kb 200 دارد که حاوی تعداد زیادی از ژن های درگیر در فرایند آلودگی است. ویژگی قابل توجه این پلاسمید این است که بخشی از مولکول DNA خود را پس از آلود کردن گیاه به درون DNA کروموزوم گیاه وارد می کند. این قطعه T-DNA نامیده می شود که بسته به سویه باکتری اندازه ای بین ۱۵ و ۳۰ کیلوباز دارد. این قطعه به شکل پایدار و به عنوان بخش کاملی از کروموزوم در سلول گیاه حفظ و به سلول های دختر منتقل می شود; اما برجسته ترین ویژگی پلاسمید Ti حضور ۸ یا تعداد بیشتری ژن در T-DNA می باشد. که این ژن ها با بیان در سلول گیاهی باعث ویژگی های سرطانی در سلول های ترانسفورم شده می شوند. این ژن همچنین ترکیبات غیر معمولی که اوپین نامیده می شوند را جهت تغذیه باکتری سنتز می کنند.
استفاده از پلاسمید Ti جهت ورود ژن های جدید به سلول گیاهی
با کشف این پلاسمید سریعا مشخص شد که می توان از آن جهت انتقال ژن های جدید به داخل سلول های گیاهی استفاده کرد. برای این کار نیاز است که ژن های جدید درون T-DNA قرار گیرند و باکتری نیز کار دشوار ورود ژن ها به DNA کروموزوم گیاهی را انجام دهد.
دو روش ابداع شده برای ورود DNA جدید به داخل پلاسمید وجود دارد:
روش وکتور دوگانه: بر اساس مشاهداتی است که نیاز به اتصال فیزیکی T-DNA به بقیه پلاسمید Ti نیست. در این سیستم دو پلاسمیدی، T-DNA روی یک مولکول نسبتا کوچک و بقیه پلاسمید به شکل طبیعی وجود دارد. در چنین روشی کارایی انتقال به گیاه به اندازه سیستم طبیعی است. در حقیقت برخی از سویه های آگروباکتریوم تومه فاشینس و آگروباکتریوم های وابسته، دارای سیستم های پلاسمیدی دوگانه هستند. پلاسمید T-DNA برای داشتن یک جایگاه محدودالاثر منحصربه فرد و دست ورزی با استفاده از تکنیک های استاندارد به اندازه کافی کوچک است.
روش هم الحاقی: با استفاده از یک پلاسمید کاملا جدید مبتنی بر وکتور E.coli که حامل بخش کوچکی از T-DNA می باشد انجام می گیرد. همولوژی بین مولکول جدید و پلاسمید Ti این امکان را فراهم می آورد که چنانچه هر دو آن ها در یک آگروباکتریوم تومه فاشینس حضور داشته باشند. نوترکیبی باعث ورود پلاسمید E.coli به محل T-DNA پلاسمید Ti شود; بنابراین، برای این کار باید ژن کلون شده درون جایگاه محدودالاثر منحصربه فردی بر روی پلاسمید کوچک E.coli الحاق شود و سپس وارد سلول های آگروباکتریوم تومه فاشینس دارای Ti گردد. فرآیند نوترکیبی باعث قرار گیری ژن جدید در درون T-DNA می شود. با آلوده کردن گیاه، ژن جدید همراه با بقیه T-DNA به درون کروموزوم های گیاهی وارد می شود.
تولید گیاهان ترانسفورم شده با پلاسمید Ti
اگر باکتری آگروباکتریوم تومه فاشینس که حاوی پلاسمید Ti دستکاری شده است، به روش طبیعی وارد گیاه شود (مثل آلوده کردن زخم روی ساقه توسط باکتری)، فقط سلول های گال تاجی حاصل دارای ژن های کلون شده خواهند بود. برای حل این مشکل چندین راه حل وجود دارد، ساده ترین راه این است که به جای آلوده کردن گیاه بالغ، کشت سلول های گیاهی یا پروتوپلاست ها را در محیط مایع آلوده کرد.
با سلول های گیاهی و پروتوپلاست هایی که دیواره سلولی آن ها دوباره شکل گرفته است، می توان مانند میکروارگانیسم ها رفتار کرد. برای مثال می توان آن ها را در محیط کشت انتخابی به منظور جدا کردن سلول های ترانسفورم شده کشت داد. گیاه بالغی که از سلول های ترانسفورم شده باززایی می شود، در هر سلول خود حاوی ژن کلون شده خواهد بود که قادر است این ژن های کلون شده را به نسل بعدی منتقل کند. باززایی گیاه ترانسفورم شده زمانی اتفاق می افتد که وکتور Ti خلع سلاح شده باشد که با این کار سلول های ترانسفورم شده خصوصیات سرطانی را نشان نمی دهند. خلع سلاح همه ی ژن های سرطانی که در T-DNA قرار گرفته اند امکان پذیر می باشد چون این ژن ها در روند آلوده سازی نقشی ندارند و آلوده سازی عمدتا توسط منطقه بیماری زایی پلاسمیدTi کنترل می شود.
پلاسمید Ri
چندسالی هست که وکتورهای کلونینگ گیاهی مبتنی بر پلاسمید Ri آگروباکتریوم ریزوژنز نیز مورد توجه قرار گرفته اند. پلاسمید های Ri و Ti بسیار شبیه به هم هستند با این تفاوت مهم که انتقال T-DNA پلاسمید Ri به جای ایجاد بیماری گال تاجی باعث بیماری ریشه مویی در گیاه می شودکه ریشه در اثر تکثیر شدید پر شاخه می شود.
کلونینگ ژن ها در گیاهان با انتقال مستقیم ژن
اگرچه پلاسمید Ti همیشه به عنوان امیدوارکننده ترین سیستم برای کلونینگ در گیاهان مطرح بوده است اما محدودیت های طیف میزبان طبیعی آگروباکتریوم تومه فاشینس سبب ترغیب جستجوی روش های جایگزین برای انتقال DNA جدید به داخل سلول های گیاه شده است که یکی از این روش ها انتقال مستقیم ژن می باشد.
انتقال مستقیم ژن به داخل هسته
در سال ۱۹۸۴ که برای اولین بار انتقال مستقیم ژن انجام شد، مشاهده شد که اگرچه پلاسمید مارپیچ مضاعف باکتریایی در سلول گیاهی قادر به همانندسازی از روی خودش نیست ولی می تواند با استفاده از نوترکیبی به یکی از کروموزوم های گیاهی وارد شود. با وجود اینکه فرآیند نوترکیبی به صورت جزئی شناخته شده است ولی به احتمال زیاد متمایز و متفاوت از فرایندهای دخیل در ورود T-DNA است.
 وکتورهای کلونینگ برای یوکاریوت ها – بخش اول (پلاسمید های الحاقی و پلاسمیدهای تکثیر شونده مخمر)
وکتورهای کلونینگ برای یوکاریوت ها – بخش اول (پلاسمید های الحاقی و پلاسمیدهای تکثیر شونده مخمر)
وکتورهای کلونینگ برای یوکاریوت ها – بخش دوم (وکتورهای کلونینگ مخمر و استفاده از کروموزوم های مصنوعی)
![]() وکتورهای کلونینگ برای یوکاریوت ها – بخش سوم (وکتورهای کلونینگ برای گیاهان عالی)
وکتورهای کلونینگ برای یوکاریوت ها – بخش سوم (وکتورهای کلونینگ برای گیاهان عالی)
نوشته شده توسط کارگروه تولید محتوا در تاریخ ۹۹/۰۲/۲۳ ساعت ۳:۳۰ ب٫ظ | تعداد دیدگاهها: ۰ دیدگاه | دستهبندی: بلاگ | برچسبها: آگروباکتریوم_تومه_فاشینس, استفاده_از_پلاسمید_Ti_جهت_ورود_ژن_های_جدید_به_سلول_گیاهی, انتقال_مستقیم_ژن_به_داخل_هسته, دوره_آموزشی_مهندسی_ژنتیک_و_کلونینگ, دوره_کارآموزی_مهندسی_ژنتیک_و_کلونینگ, راهنمای_آموزشی_وکتورهای_کلونینگ_برای_گیاهان_عالی, رفع_اشکالات_وکتورهای_کلونینگ_برای_گیاهان_عالی, روش_ورود_DNA_جدید_به_داخل_پلاسمید, فایل_pdf_وکتورهای_کلونینگ_برای_گیاهان_عالی, مبانی_و_مفاهیم_وکتورهای_کلونینگ_برای_گیاهان_عالی, وکتورهای_کلونینگ_برای_گیاهان_عالی, پلاسمید_Ri, کارگاه_آموزشی_پذیرش_تحصیلی_علوم_زیستی